Introduction
Lumen are cavities that extend into the body.
Water accounts for 50-70% of body weight. Water content of the body correlates inversely with fat content.
There are four main fluid compartments:
- Intracellular fluid inside of cells
- Interstitial fluid outside of cells and blood vessels
- Plasma inside of blood vessels
- Glycocalyx
Homeostasis
Homeostasis is the ability to keep the internal environment within a tolerable range.
Feedback systems maintain homeostasis. In a positive feedback system, stimuli is received by the receptor, which sends the information to the integrating center, which sends instructions to the effector, which carries out the response. For example, a drop in blood pressure causes the heart to speed up and the kidneys to retain fluid.
Cell Physiology
Mitochondria converts sugar, fats, proteins, and O2 into ATP and CO2 through cellular respiration.
The cell membrane is a protein-studded phospholipid bilayer. The head of the phospholipid is polar (hydrophilic) and the fatty tails are nonpolar (hydrophobic).
The membrane is selectively permeable, allowing some substances to pass while blocking others. Small, nonpolar molecules (O2, CO2, ethanol) can pass through easily. Large polar molecules (glucose) and ions (Na+, K+) require assistance.
Oxygen and nitrogen are highly electronegative, causing them to pull electrons toward themselves in a covalent bond. This creates a polar molecule with partial positive and negative charges.
Proteins
Amino acids can link together to form:
- Peptides (2-9 amino acids)
- Polypeptides (10-100 amino acids)
- Proteins (>100 amino acids)
The structure of proteins can be described in four levels:
- 1st structure is the sequence of amino acids
- 2nd structure is folding into α-helixes and β-pleated sheets
- 3rd structure is the overall fibrous or globular shape
- 4th structure is the combination of multiple proteins
There are four forces that fold a polypeptide chain:
- Hydrogen bonds form between polar groups
- van der Waals forces form from temporary dipoles
- Ionic bonds form between charged groups
- Covalent bonds form from sharing electrons
Hemoglobin is dull red when deoxygenated and bright red when oxygenated. This is an example of how conformational changes in proteins can affect their function.
Cell Transport
Molecules can get across the membrane in four ways:
- Diffusion
- Facilitated diffusion (ion channels, carrier proteins)
- Active transport (primary, secondary, vesicular)
Diffusion
The rate of diffusion, or flux, is affected by
- Solubility of molecule ()
- Surface Area ()
- Concentration gradient ()
- Size of molecule ()
- Distance ()
- Temperature
This is given by the equation:
Sometimes the solubility, size of the molecule, and distance are combined into a permeability coefficient ():
Permeability is affected by several factors:
- Non-polar molecules are more permeable than polar molecules
- Weak acids are permeable when protonated
- Weak bases are permeable when unprotonated
Ion Trapping
Suppose there is a pH gradient across the membrane. Then more weak acids will protonated into on the side that has a higher concentration of . This will increase diffusion of across the membrane. Once inside the cell, the weak acid will dissociate again, trapping it inside.
Note that . That means that a lower pH corresponds to a higher concentration of ions.
Drugs are usually weak acids or weak bases, so they can take advantage of ion trapping. Pharmaceutical companies designing drugs must take into account that the pH of the stomach is between 1-2, and the pH of the small intestine is between 6-7.
Facilitated Diffusion
Facilitated diffusion can happen through ion channels and carrier proteins.
Ion channels have three properties: selectivity, permeability, and gating. Outside of the cell, each ion is surrounded by water molecules (sphere of hydration), which gives the ion some geometry. The aqueous pore of the ion channel has the same geometry as the sphere of hydration, allowing the ion to pass through.
Carrier proteins have three properties: specificity, saturability and competition. Saturability and competition arise because there are a limited number of carrier proteins and potentially multiple types of molecules that can bind to them.
Here are the intracellular and extracellular concentrations of ions:
| Ion | Intracellular (mM) | Extracellular (mM) |
|---|---|---|
| Na+ | 15 | 142 |
| K+ | 150 | 4 |
| Cl- | 5 | 120 |
| Ca++ | 0.0001 | 1 |
Active Transport
Primary active transport uses ATP to move molecules against their concentration gradient.
An example is the sodium-potassium pump (Na+/K+-ATPase), which moves 3 Na+ ions out of the cell and 2 K+ ions into the cell for each ATP hydrolyzed.
Secondary active transport uses the energy from one molecule moving down its concentration gradient to move another molecule against its concentration gradient. This can be done via cotransport (same direction) or countertransport (opposite direction).
For example, the sodium–glucose cotransporter (SGLT) uses the energy of Na+ moving down its electrochemical gradient to transport glucose into the cell against its concentration gradient. This process is driven in part by the negative resting membrane potential.
Vesicular transport uses vesicles to move large molecules across the membrane. Endocytosis brings molecules into the cell, while exocytosis sends molecules out of the cell.
Osmosis and Tonicity
Osmosis
Osmosis is the diffusion of water across a selectively permeable membrane (through aquaporin channels).
Osmotic pressure () is a measure of how strongly water wants to move into a solution. If one side of the membrane has more solute particles (and higher osmotic pressure), then water will move toward that side.
The equation for osmotic pressure is given by:
Where:
- is the concentration of solute particles in mOsm/L
- is the temperature in K
- is the gas constant in atm/mol · K
Note that mOsm is the number of particles a substance dissolves into in the solution. For example, NaCl dissolves into Na+ and Cl-, so 1 mole of NaCl produces 2 Osm.
Tonicity
Tonicity is a measure of how a solution affects cell volume.
- Hypertonic solutions cause cells to shrink
- Isotonic solutions cause no change in cell volume
- Hypotonic solutions cause cells to swell
For human cells, an isotonic solution is generally 300 mOsm/L.
Note that tonicity is only affected by the concentration of non-penetrating solutes. There is also a measure called osmolarity that depends on the total concentration of solutes (both penetrating and non-penetrating).
Filtration and Absorption
Fluid moves from the capillaries into the interstitial fluid (filtration), and from the interstitial fluid back into the capillaries (absorption). To do this, it must pass through the endothelial cells that line the capillaries. This can happen through diffusion, bulk flow, or vesicular transport.
Fluid exchange between capillaries (plasma) and interstitial fluid is governed by Starling forces:
- Hydrostatic pressure caused by the pumping action of the heart pushes fluid out of the capillaries. This pressure decreases along the length of the capillary.
- Colloid osmotic pressure from the proteins within the capillary pulls fluid into the capillaries. It remains relatively constant along the length of the capillary.
This is why filtration generally occurs at the arterial end of the capillary (higher hydrostatic pressure) and absorption occurs at the venous end of the capillary (lower hydrostatic pressure).
Starling forces also explains why elevating an injured limb helps reduce swelling. It decreases hydrostatic pressure, reducing fluid leakage into the interstitial space.
Recently, the glycocalyx layer lining the endothelial cells has been found to play an important role in fluid exchange. It acts as a barrier to protein movement, modulating colloid osmotic pressure across the capilary. This promotes more filtration and less absorption.

Cystic Fibrosis
Cystic fibrosis is a genetic disorder caused by mutations in the CFTR gene, which encodes a chloride channel protein. Cystic fibrosis causes the chloride channel to be malformed and marked by degradation, preventing it from reaching the cell membrane.
Chloride is trapped inside of the cell, preventing water from moving outside to thin the mucus. Chloride is also not reabsorbed, resulting in very salty sweat.
In the pancreas, the lack of chloride secretion limits Na+ secretion, which then limits water secretion, causing pancreatic exocrine secretion to be too thick. Finally, this causes digestive enzymes to not be secreted into the GI track.
Some treatments for cystic fibrosis include:
- Drugs that chaperone CTFR to the membrane to prevent its premature degradation
- Drugs that increase conductance of Fl
Cell Signaling
Chemical communication occurs in the nervous system and the endocrine system.
Steps:
- Ligand released
- Ligand binds to receptor
- Receptor protein have conformational change
Receptors differ in the number of ligands they can bind to (specificity) and their affinity for those ligands.
If there are multiple types of ligands that can bind to the same receptor, then they will compete for binding. Some ligands are agonists, which activate the receptor, while others are antagonists, which block the receptor. Receptors can become saturated when all of the receptors are occupied by ligands.

There are serveral types of receptor responses:
- Ligand-gated ion channels open or close
- Tyrosine kinase have enzymatic activity
- Receptors interact with cytoplasmic janus kinases
- G-protein coupled receptors activate
G-proteins contain an alpha, beta, and gamma subunit. When the first messenger binds to the receptor, it increases the affinity of the alpha subunit for guanosine triphosphate (GTP). When bound to GTP, the alpha subunit dissociates and links up with an effector protein in the membrane. Once it activates the protein, the GTP is cleaved back into GDP, allowing the alpha subunit to recombine with the G-protein group.
When adenylyl cyclase is activated by the alpha subunit, it takes in ATP and creates cyclic adenosine monophosphate (cAMP). cAMP acts as a second messenger and binds to an enzyme known as cAMP-dependent protein kinase (protein kinase A). Eventually, cAMP terminates when it is broken down to AMP.
Question: Why is this process so complicated?
The benefits of having a multistep process is (1) amplification, (2) modulation, and (3) duration.

Question: How can the same second messenger cause two different cellular responses?
Each receptor is surrounded by different effector proteins, protein kinases, and phosphatases. This allows the same second messenger to cause different responses in different compartments.
Endocrine System
The hypothalamus produces releasing factors that are released into the capillary bed in the pituitary stalk. They stimulate the anterior pituitary gland, causing it to release hormones through the bloodstream. These hormones can have direct effects on target tissues or stimulate other endocrine glands to release hormones.
For example, the hypothalamus releases thyrotropin-releasing hormone (TRH), which stimulates the anterior pituitary to release thyroid-stimulating hormone (TSH). TSH then stimulates the thyroid gland to release thyroid hormones (T3 and T4) in the bloodstream. T3 and T4 are lipid-soluble hormones that enter cells and bind to nuclear receptors, affecting gene transcription.
Hyperthyroidism causes increased metabolic rate and body temperature.
Question: How do you store something that is lipid-soluble?
T3 and T4 are covalently bonded together in a non-penentrating protein called thyroglobulin and stored in the colloid of the thyroid gland. When T3 and T4 are needed, the cell endsocytoses the thyroglobulin, breaks the covalent bond, and releases T3 and T4 into the bloodstream.
Question: What is determining the volume of the thyroid gland?
Most of the volume of the thyroid gland comes from the colloid. When the colloid swells up, the thyroid gland swells up and becomes a goiter. The colloid swells up when there is high osmotic pressure (caused by a high concentration of thyroglobulin).
Question: What happens when...
| Condition | TSH | T3 & T4 | Goiter |
|---|---|---|---|
| Hyperactive thyroid stimulation | Low | High | Yes |
| Hyperactive hypothalamus or anterior pituitary | High | High | Yes |
| Hypersecreting thyroid | Low | High | No |
| Thyroid failure or low iodine | High | Low | Yes |
| Inactive hypothalamus or anterior pituitary | Low | Low | No |
Neurons, Synapses, and Signaling
Membrane Potential
All cells exhibit a membrane potential (Vm) across the membrane. The inside of the cell is typically negative relative to the outside. This potential energy can be used to do work. Excitable cells (neurons, muscles) use changes in membrane potential to send signals.
Cells use energy (ATP) to create a chemical gradient with K+ and Na+. K+ has a higher concentration inside the cell, while Na+ has a higher concentration outside the cell.
If more K+ channels are opened, then more K+ will leave the cell, making the inside more negative. If more Na+ channels are opened, then more Na+ will enter the cell, making the inside more positive. Ion channels will not appreciably change the concentration of K+ and Na+.
Note that 3 Na+ are pumped out of the cell and 2 K+ are pumped into the cell for each ATP hydrolyzed, so there is also a small membrane potential from that.
If there are multiple permeable ions, then the membrane potential is given by the Goldman-Hodgkin-Katz equation:
Where:
- is the permeability of the ion, which is impacted by number of ion channels
- is the concentration of the ion
Equilibrium potential
To reach electrochemical equilibrium, the chemical gradient must be balanced by the electrical gradient. Relatively few ions need to move to create a significant electrical gradient. This is described by the Nernst equation:
Where:
- is the gas constant
- is the temperature
- is the ion charge
- is Faraday's constant
Current
From Ohm's Law, we can derive this equation for current:
Where:
- is current
- is the membrane conductance
- is the membrane potential
- is the equilibrium potential
Note that:
- When is positive, there is an outward current (positive charge exiting the cell or negative charge entering the cell).
- is maintained at a constant level.
- changes rapidly.
- has some components that are constant (leak channels) and some components that change rapidly (gated channels). can change rapidly.
- Driving force is the difference between the membrane potential and the equilibrium potential .
If the number of open ion channels changes, then the conductance changes, which changes the current . This causes more particles to either enter or leave the cell, which changes membrane potential . Membrane potential asymptote toward equilibrium potential . At steady-state, the net current should be zero.
If there is a delayed rectifier potassium channel, the membrane potential may undershoot the equilibrium potential.
Neurons
Neurons are very large cells with high metabolic demands. As soon as resources are cut off, they immediately start to malfunction. Long projecting neurons are the first to show signs of disease, which manifests as tingling or numbness in hands in feet.
Astrocytes line the capillaries and tightly regulate what can come out of it (blood-brain barrier).
Afferent neurons receive signals from sensory receptors. Interneurons in the central nervous system connect the afferent neurons to the efferent neurons. The efferent neuron sends signals to muscles, glands, and other tissue through the axon terminal.
Neuron anatomy:
- Dendrites receive inputs from other cells
- Soma (cell body) integrates all input
- Axon hillock decides whether to fire an action potential
- Axon conducts the action potential down the length of the neuron
- Presynaptic axon terminals release chemical signalling molecules
Action Potential
Neurons communicate with each other through action potentials and neurotransmitters.
- An action potential arrives at the presynaptic axon terminal.
- The depolarization causes voltage-gated calcium channels to open.
- Ca2+ enters the cell, causing exocytosis of synaptic vesicles containing neurotransmitters.
- The neurotransmitters diffuse across the synaptic cleft and bind to receptors on the postsynaptic cell.

Voltage-gated sodium channels have three states: closed, open, and inactivated. When depolarizing stimulus brings the membrane to a threshold potential, the voltage-gated sodium channels rapidly open, allowing Na⁺ to enter the cell and further depolarize the membrane. This depolarization triggers additional sodium channels to open through positive feedback, creating the action potential. Shortly after opening, the channel becomes inactivated, preventing further Na⁺ influx even if the membrane remains depolarized. The channel cannot reopen until the membrane repolarizes toward its resting potential, at which point it returns to the closed state.
Delayed rectifier potassium channels are voltage-gated channels that open in response to depolarization. As the voltage-gated sodium channels are inactivated, the potassium channels open up, allowing the membrane to return back toward its resting potential. This is an example of a negative feedback loop.

Conduction and Synapses
Graded potentials can only travel a short distance before diminishing in amplitude. To send signals beyond a very short distance, action potentials must be generated and propagated along the entire length of neuron.
Current enters the cell and has three pathways:
- Down the inside of the axon ()
- Through the cell membrane ()
- To the cell membrane to charge up the capacitance ()
We would like current to travel down the axon, but the problem is that axons are poor conductors ( is high), and pathways (2) and (3) are significant drains on overall current.
These problems are solved by several adaptations:
- Having larger axon diameters, decreasing
- Having no open ion channels between nodes, increasing
- Having a thick myelin sheath, decreasing
Every so often, there are gaps in the myelin sheath called nodes of Ranvier where the voltage-gated sodium channels are concentrated. These regions are where the action potential is regenerated.
Local anesthetic works by blocking action potentials in small diameter axons (like those that handle pain). It spares large diameter axons like touch and motor.
Nervous System
Types of Nervous Systems
The central nervous system (CNS) is comprised of the brain and spinal cord.
- The hypothalamus controls the pituitary hormones.
- The medulla controls breathing and heart rate.
The peripheral nervous system (PNS) has three components:
- The somatic controls voluntary movements by skeletal muscles.
- The autonomic controls involuntary functions by smooth muscles and glands.
- The enteric controls the gastrointestinal system (gut).
The autonomic nervous system can be divided into the sympathetic, which is responsible for the fight-or-flight response, and the parasympathetic, which is responsible for rest-and-digest functions.
Activation Pathways
In the somatic nervous system, the axon releases acetylcholine (ACh), which binds to nicotinic ACh receptors on the skeletal muscle, causing it to contract.
In the automatic nervous system, preganglionic neurons releases ACh, which binds to nicotinic ACh receptors on the postganglionic neuron. The postganglionic then releases a different neurotransmitter depending on whether it is part of the sympathetic or parasympathetic nervous system.
In the sympathetic nervous system, the postganglionic neuron then releases norepinephrine, which binds to adrenergic receptors on the target tissue. The adrenergic receptors can be either alpha or beta, and they can have different effects on the target tissue. Alpha-1 adrenergic receptors cause vasoconstriction, while beta-2 adrenergic receptors cause vasodilation.
In the parasympathetic nervous system, the postganglionic neuron then releases ACh, which binds to muscarinic ACh receptors on the target tissue
Visceral and Somatic Motor System
Muscles are made of muscle fibers, which contain myofibrils composed of repeating sarcomeres. The sarcomere is the basic contractile unit of muscle and contains thin filaments (actin) anchored to the zig-zagging Z lines and thick filaments (myosin) anchored at the middle M line. The actin contains regulatory proteins (troponin and tropomyosin) which control whether it can bind to myosin.

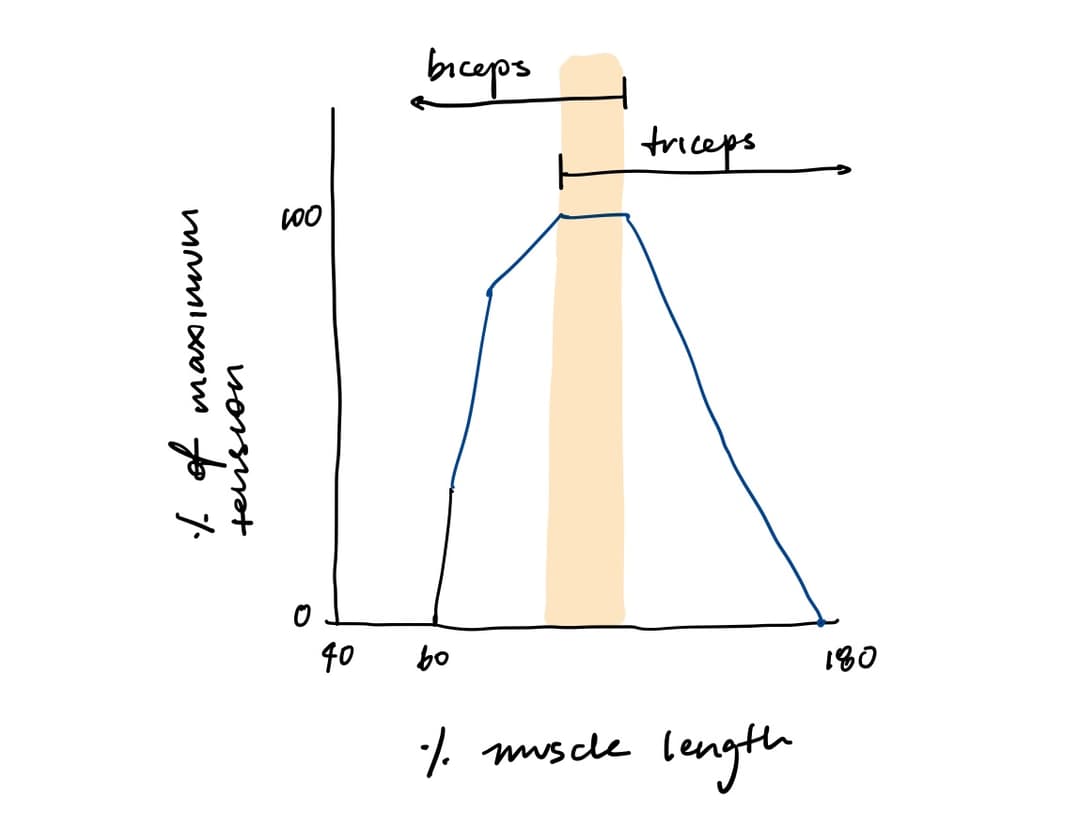
Sarcomeres have an optimal length at which they can generate the most force. If they are stretched too much, then there will be less overlap between actin and myosin, so fewer cross-bridges can form. If they are stretched too little, then actin filaments from one side interfere with crossbridge formation on the other side. This is called the length-tension relationship.
For example, when the arm is extended, both biceps and triceps have optimal overlay. When the arm is flexed, biceps are shorter than optimal and triceps are longer than optimal.
The neuromuscular junction is the synapse between a motor neuron and a skeletal muscle fiber. The motor neuron releases ACh, which binds to nicotinic ACh receptors on the muscle fiber, causing depolarization of the sarcolemma and propagation of an action potential.
T-tubules conduct the action potentials, allowing them to reach the interior of the cell. In the T-tubules, depolarization activates dihydropyridine (DHP) receptors. They are mechanically coupled to ryanodine receptors on the sarcoplasmic reticulum, so their conformational change causes Ca++ to release from the sarcoplasmic reticulum.
Calcium binds to troponin, causing tropomyosin to shift away from myosin-binding sites on actin. A crossbridge forms between the myosin heads and the actin when myosin has ADP and Pi bound and troponin has Ca++ bound. This causes muscle contraction.
Muscular force is correlated with (1) the concentration of intracellular calcium as well (2) the number of cross-bridges per second.
Cardiac System
The heart is a dual pump:
- The pulmonary side pumps blood through the pulmonary artery to the lungs
- The systemic side pumps blood through the aorta to the rest of the body
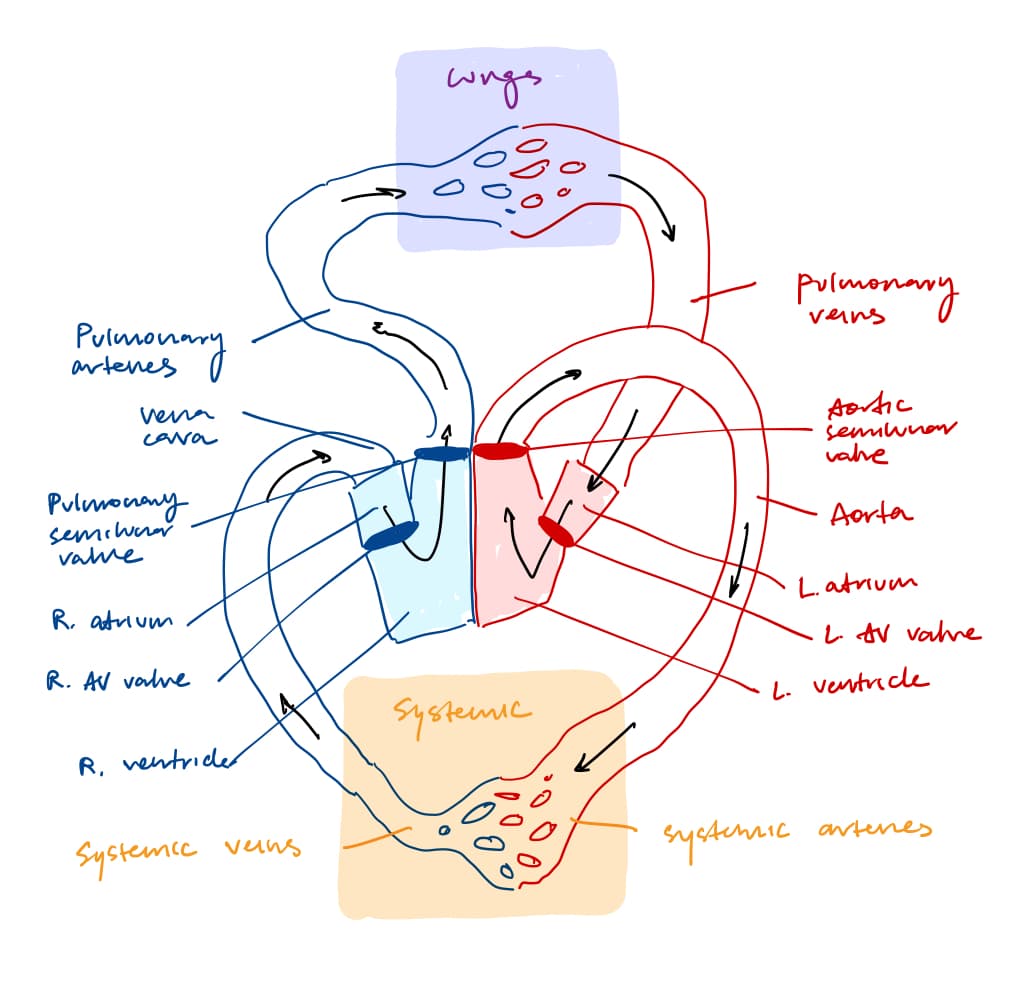
The atrium receives blood moving into the heart. The ventricle receives blood from the atrium.
Valves open and close depending on pressure in the two chambers. Valves can be blown open (prolapse) if the pressure difference is too high, leading to backflow of blood. Capillary muscles hold the valve in place to prevent a prolapse.
There are multiple valves in the heart:
- The atrium-ventricular (AV) valve separates atrium and ventricle
- The aortic and pulmonary semilunar valves separate the ventricles from the aorta and pulmonary artery
Usually, the first sound of the heartbeat is caused by the closing of the AV valves, and the second sound is caused by the closing of the semilunar valves. Murmurs can be caused by valves not opening well (stenoic) or not closing well (insufficient).
Arteries bring blood away from the heart and veins bring blood toward the blood. Capillary beds are where the blood becomes deoxygenated and picks up CO2.
Cardiac Excitation
Pacemaker cells in the sinoatrial (SA) node automatically generate heartbeats and pass signals to contractile muscle cells through gap junctions.
- When the membrane is very negative, funny NA+ channels open; Na+ enter the cell
- Voltage-gated T-type Ca2+ channels to open briefly; Ca2+ enter the cell
- At threshold, L-type Ca2+ channels open; more Ca2+ enter the cell
- The action potentials move to the contractile cells, causing them to contract and pump blood
The SA node is the main pacemaker and operates at 60-100 bpm. The atrioventricular (AV) node can also act as a pacemaker, but it generates action potentials at a slower rate of 40-60 bpm. If both the SA and AV nodes fail, the His bundle can generate action potentials at an even slower rate of 20-40 beats per minute.
From the SA node, the action potential travels from the atria to the AV nodes to the His bundle to to the ventricles.
Ventricular action potential:
- Rapid depolarization: voltage-gated Na+ channels open; Na+ enter the cell
- Brief repolarization: voltage-gated Na+ channels inactivate; some K+ channels open; K+ exit the cell
- Plateau: voltage-gated Ca2+ channels open; Ca2+ enter the cell and balance the outward K+ current, creating a plateau in membrane potential
- Repolarization: voltage-gated Ca2+ channels inactivate, and more K+ channels open, allowing K+ to exit the cell and repolarize the membrane back to its resting potential
EKG
A EKG consists of a few phases:
- P wave: depolarization of the atria
- PR interval: conduction delay in the AV node
- QRS complex: depolarization of the ventricles
- T wave: repolarization of the ventricles
Cardiac Cycle
In a cardiac cycle, there is first isovolumetric relaxation, where both valves are closed as pressure rises. Then the AV valves open, the semilunar valves close, and blood fills the ventricles (diastole). Then there is isovolumetric contraction again, where both valves are closed as pressure rises. Then the AV valves close, the semilunar valves open, and blood is ejected (systole).
| Term | Definition |
|---|---|
| Diastole | Muscle relaxation in ventricles |
| Systole | Muscle contraction in ventricles |
| Isovolumetric contraction | Period where both valves are closed as pressure rises |
| End diastolic volume | Volume of blood in ventricles at the end of diastole (~120 mL) |
| End systolic volume | Volume of blood in ventricles at the end of systole (~50 mL) |
| Stroke volume | Amount of blood ejected from ventricles (EDV - ESV) |
| Ejection fraction | Percentage of blood ejected from ventricles (SV/ EDV) |
| Preload | End-diastolic volume |
| Afterload | Resistance ventricles must overcome to eject blood |
Frank-Starling Mechanism
The Frank-Starling mechanism states that the more the ventricles are filled during diastole, the more they will contract during systole.
Question: There is no change in sympathetic and parasympathetic tones, so it must be an intrinsic property of the heart. How does it work?
Sarcomeres can generate the most force when they are stretched to 2.0-2.2 micrometers. When the ventricles are filled more, the sarcomeres are stretched closer to their optimal length, allowing for more cross-bridges to form and a stronger contraction.
The problem is that the length-tension effect only accounts for a small increase in force. There are two additional mechanisms that contribute to the Frank-Starling mechanism:
- Stretch-sensitive calcium channels let more calcium into the cell, allowing more cross-bridges to form.
- Stretch-sensitive troponin binds more strongly to calcium, allowing more cross-bridges to form.
Increased sympathetic stimulation can also cause increased calcium concentration and increased force.
Pressure-Volume Loops
The PV loop represents the changes in pressure and volume in the ventricles during a single cardiac cycle.
There are four phases of the PV loop:
- Isovolumetric relaxation
- Diastolic filling
- Isovolumetric contraction
- Systolic ejection
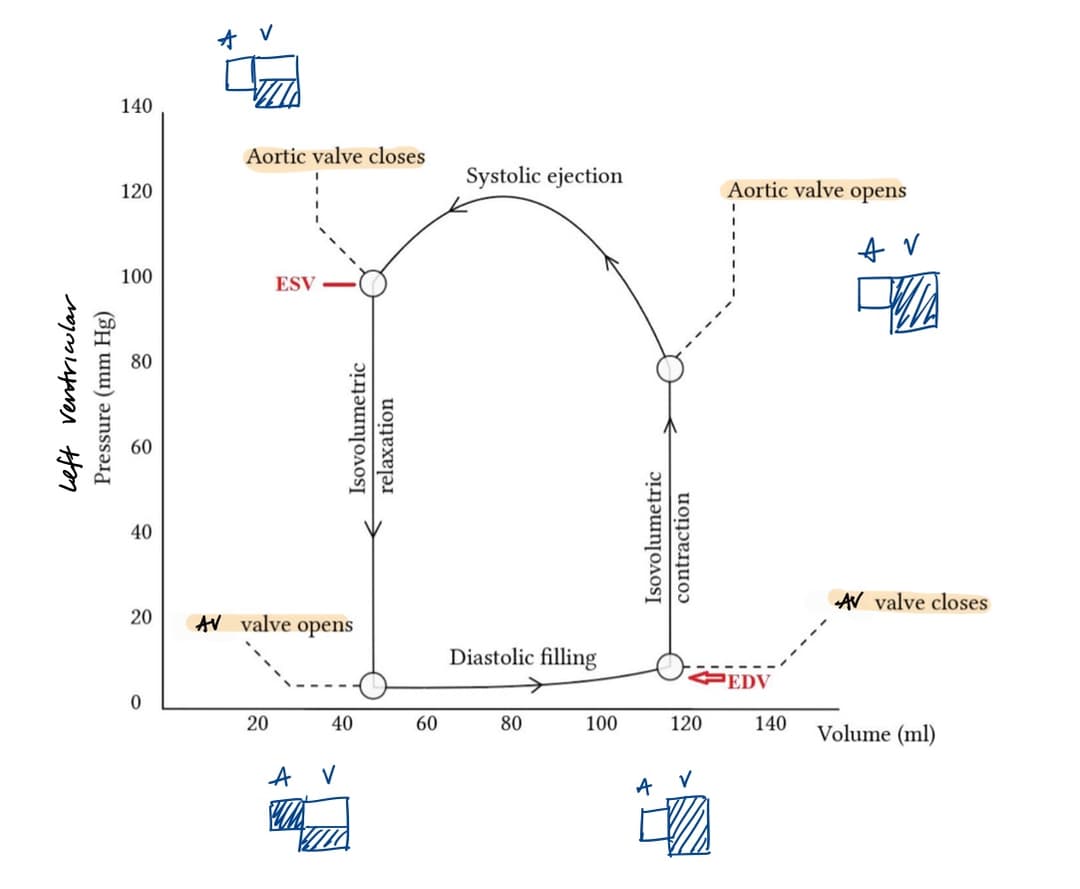
The diastolic pressure curve represents the passive filling of the ventricle and is impacted by how easily the ventricle stretches (compliance).
The systolic pressure curve represents the resistance the ventricle must overcome to eject blood and is impacted by how strongly the ventricle contracts (contractility).
Contractility is influenced by the Frank-Starling mechanism, which is affected by preload, afterload, and sympathetic tone.
The actual preload is determined by the venous return, and the actual afterload is determined by the arterial pressure and TPR.
Stroke volume is impacted by preload, afterload, and sympathetic tone. If preload is higher, then the Frank-Starling mechanism kicks in—resulting in higher stroke volume. If afterload is lower, then more work can go into ejecting volume rather than generating higher pressure—resulting in an increase in stroke volume.
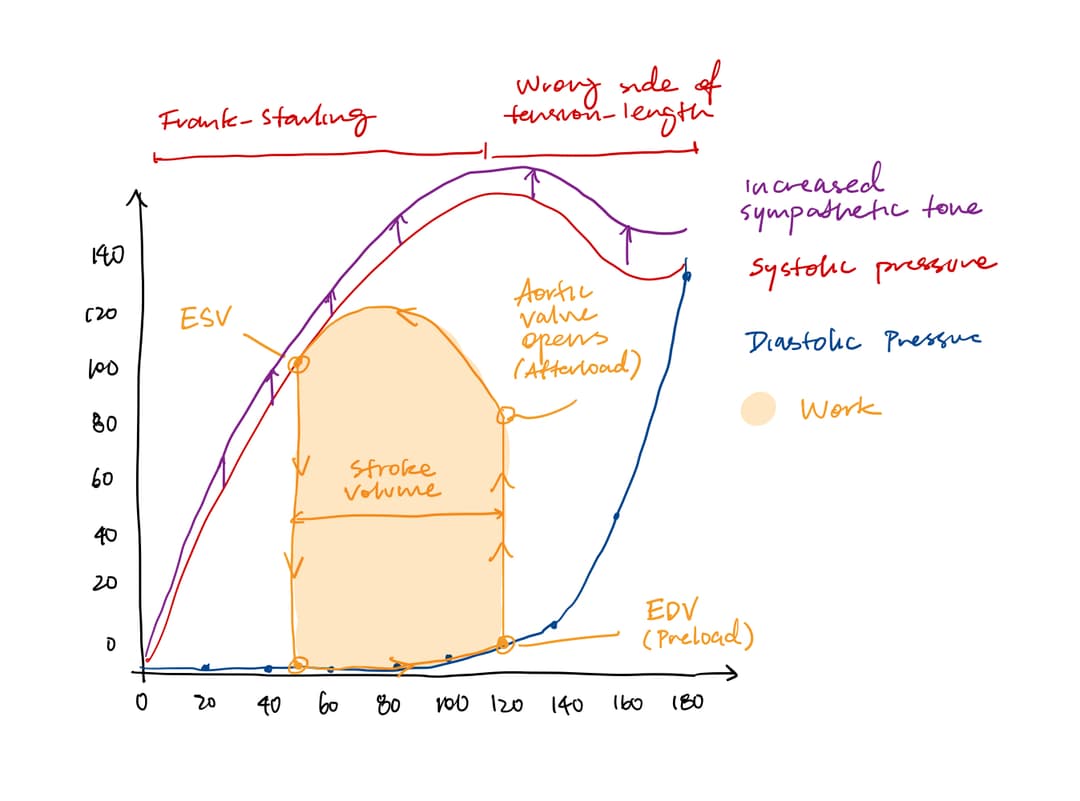
Wiggers Diagram
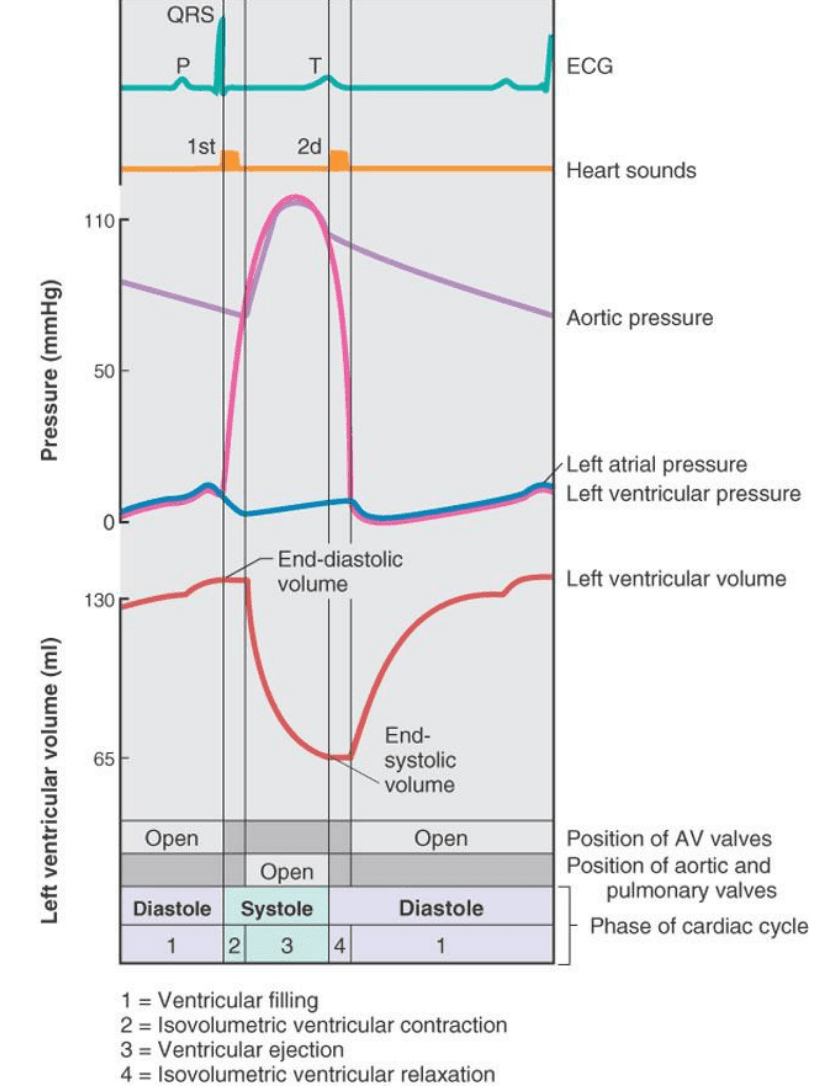
Notes:
- Whenever the pressure curves cross, a valve changes state
- Most filling of the ventricles is passive
- Most filling ocurrs in the first third of diastole
- Most emptying occurs in the first third of systole
Blood Flow
Blood Cells
Blood cells don't have a nucleus, so they can't make new proteins. They have a lifespan of around 120 days, after which they are removed by the spleen and replaced by new blood cells made in the bone marrow.
RC Circuit
Blood flow is similar to an RC circuit. The small blood vessels have high resistance. The arteries have low resistance and act as a capacitor for pressure.
As you age, your arteries become stiffer, so they are less able to stretch and recoil. This causes a higher systolic pressure and a lower diastolic pressure, leading to a higher pulse pressure.
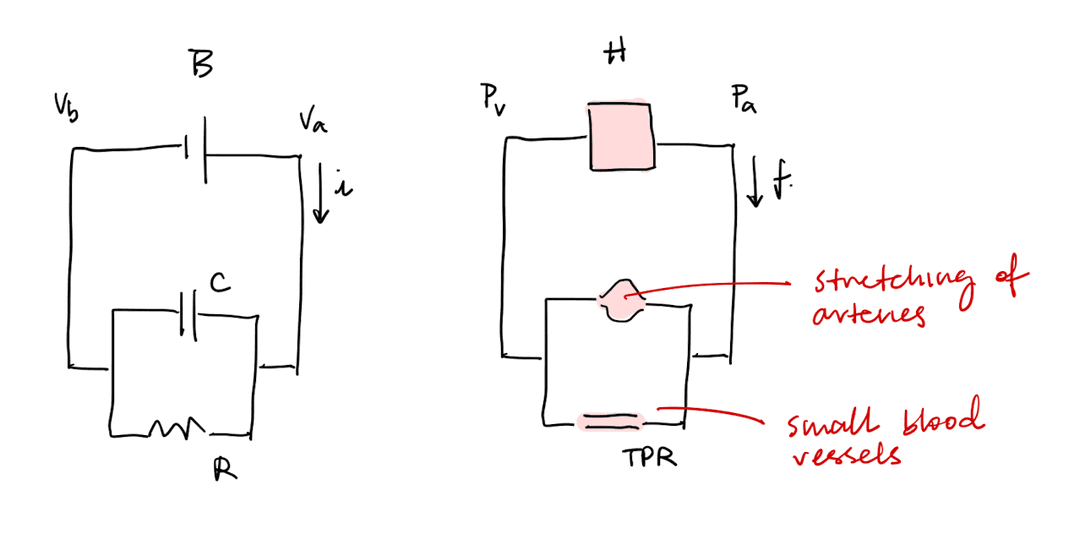
Increased capacitance of the veins decreases venous return, decreasing preload and thus stroke volume. Increase capacitance of arteries decreases the amount that pressure rises when the heart ejects blood (pulse pressure). Note that veins tend to have greater capacitance than wrteries.
Capacitance of the veins and arteries does not affect total peripheral resistance, which is determined by the arterioles.
Blood Pressure
The equation for blood pressure is
Where:
- is arterial pressure
- is venous pressure, which is almost
- CO is cardiac output
- TPR is total peripheral resistance
The equation for cardiac output is
Where:
- CO is cardiac output (mL/min)
- HR is heart rate (beats/min)
- SV is stroke volume (mL/beat)
Heart rate in the SA node is determined by the degree of sympathetic and parasympathetic stimulation. Stroke volume is determined by contractility and the Frank-Starling mechanism, which is influenced by preload afterload, and sympathetic tone. One tricky part is that affects the afterload, which then affects stroke volume. When increases, the afterload increases, which decreases stroke volume.
Total peripheral resistance is the overall resistances in the blood vessels. Resistance of the blood vessels is determined by Pouseille's Law, which states
Where:
- is resistance
- is the viscosity of the fluid
- is the radius of the vessel
TPR is only affected by vasodilation and vasoconstriction, which is impacted by sympathetic and parasympathetic tone.
Pressure Gradient
Blood pressure is highest in the aorta and decreases as it moves through the arteries, arterioles, capillaries, venules, veins, and back to the heart. The largest drop in pressure occurs in the arterioles, which are the main site of resistance in the circulatory system.
Average aortic pressure is diastolic pressure plus one-third of the pulse pressure, or around 100 mmHg.
Question: Blood flows out of the heart because there is a standing pressure gradient. But there is no pressure gradient to drive blood flow in the veins. How does blood flow back to the heart?
There is a lot of blood contained in chambers separated by valves. When joints and muscles push on the walls of these chambers, pressure increases, forcing blood to flow through the one-way valves back to the heart.
Actually, if you stand still for a very long time, there may not be enough blood flowing back to the heart, causing you to faint.
Sympathetic and Parasympathetic Tone
The brain monitors and maintains blood pressure through baroreceptors in the carotid sinus and aortic arch. When their action potential firing decreases, the brain increases sympathetic and decreases parasympathetic activity to raise blood pressure.
For example, during a valsalva maneuver, you exhale forcefully against a closed airway, which increases pressure in the chest and decreases venous return. This causes a decrease in stroke volume and cardiac output, which causes a drop in blood pressure. The baroreceptors detect this drop in blood pressure and increase sympathetic tone to compensate.
Effects of sympathetic tone:
- Contractility increases, causing the heart to beat faster and stronger. This increases stroke volume and cardiac output.
- Vasoconstriction increases, causing total peripheral resistance to increase. This helps maintain blood pressure even when cardiac output increases.
- Veins stiffen, decreasing capacitance. This causes a greater increase in pressure when the muscles compress the veins, resulting in a greater venous return. As a result, preload increases, which increases stroke volume and cardiac output.
Specifically, the sympathetic system uses norepinephrine to bind to alpha-1 adrenergic receptors on the blood vessels, causing vasoconstriction and increasing TPR. It also uses norepinephrine to bind to beta-1 adrenergic receptors on the heart, increasing heart rate and contractility, which increases cardiac output.
The parasympathetic system uses ACh to bind to muscarinic receptors on the heart, decreasing heart rate and contractility, which decreases cardiac output.
Vasodilation and Vasoconstriction
Smooth muscle and precapillary sphincter muscles are found in the walls of blood vessels. They can contract or relax to change the radius of the vessel, which changes resistance and blood flow.
Precapillary sphincter muscles regulate their own oxygen supply. When they constrict, less blood flows to local capillaries and oxygen levels drop. When oxygen levels drop, they can no longer manufacture ATP at the same rate, causing them to become weak and relaxed.
The strength of the precapillary sphincter muscles is determined by the number of crossbridges formed, which is affected by the concentration of calcium and the ATP.
The following factors can cause dilation:
- Decreased oxygen levels
- Increased temperature
- Increased carbon dioxide levels
- Decreased pH
- Increased adenosine levels
Multiple factors can affect blood flow:
- Precapillary sphincter muscles
- Upstream artery effect through nitric oxide
- Hormones in the blood such as histamine, epinephrine, antidiuretic hormone (ADH) and angiotensin
Vasodilation in the precapillary sphincter muscles causes an increase in blood flow, which increases shear forces on the upstream arteries. This causes an increase in nitric oxide production, which causes vasodilation in the upstream arteries. This is called the upstream artery effect.
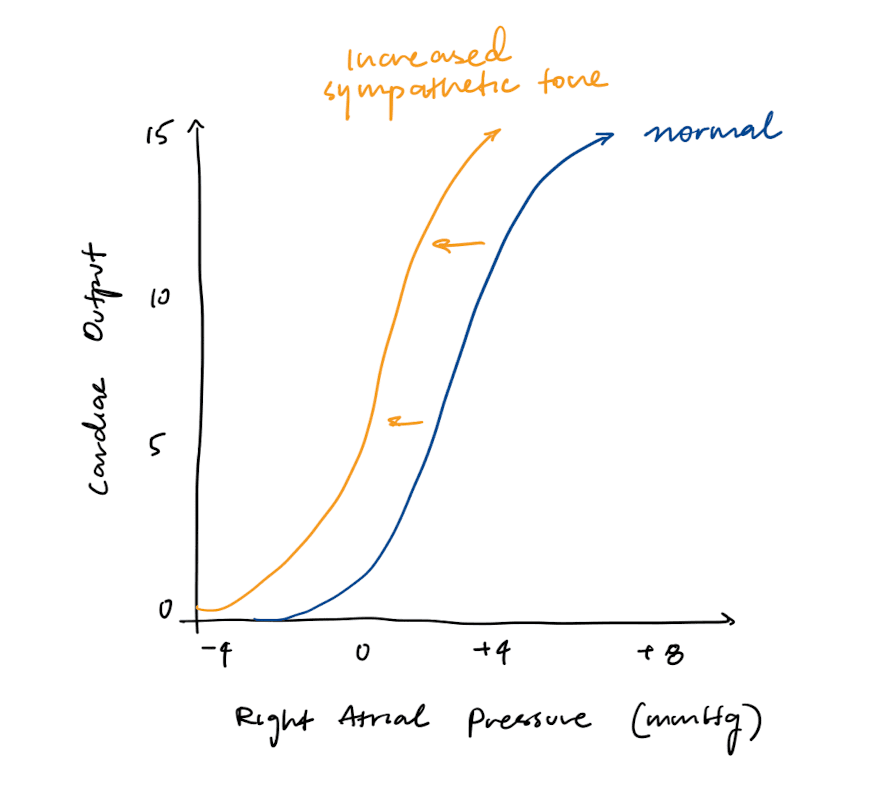
Cardiac Function Curve
The cardiac function curve shows that as right atrial pressure (RAP) increases, there is more filling (preload), increasing cardiac output.
High right atrial pressure implies that there is a high venous return. Assuming that the right heart pushes blood effectively through the pulmonary system to the left heart, this means that there is high preload in the left ventricles as well. High preload causes increased stroke volume and thus increased cardiac output.
There are two things that can affect the cardiac function curve:
- Heart rate
- Contractility
Increasing sympathetic tone increases both heart rate and contractility, so it will shift the cardiac function curve to the left.
Vascular Function Curve
The vascular function curve shows that as right atrial pressure (RAP) increases, there is less of a pressure gradient, decreasing venous return.
Blood flow between the veins and the right atrium depends on the pressure gradient between them. When right atrial pressure is high, there is a smaller pressure gradient, so blood flows back to the heart slowly, decreasing venous return.
There are three things that can affect the vascular function curve:
- Blood volume
- Capacitance of veins
- Total peripheral resistance.
If blood volume or capacitance of veins increases, then the venous return will increase at a given right atrial pressure, shifting the vascular function curve to the right.
Mean circulatory filling pressure (MCFP) is the pressure within the entire cardiovascular system when blood flow ceases. It is represented by the x-intercept, which shifts to the right when blood volume or capacitance increases.
If total peripheral resistance increases, then the venous return will decrease multiplicatively at every right atrial pressure. The slope of the vascular function curve will decrease.
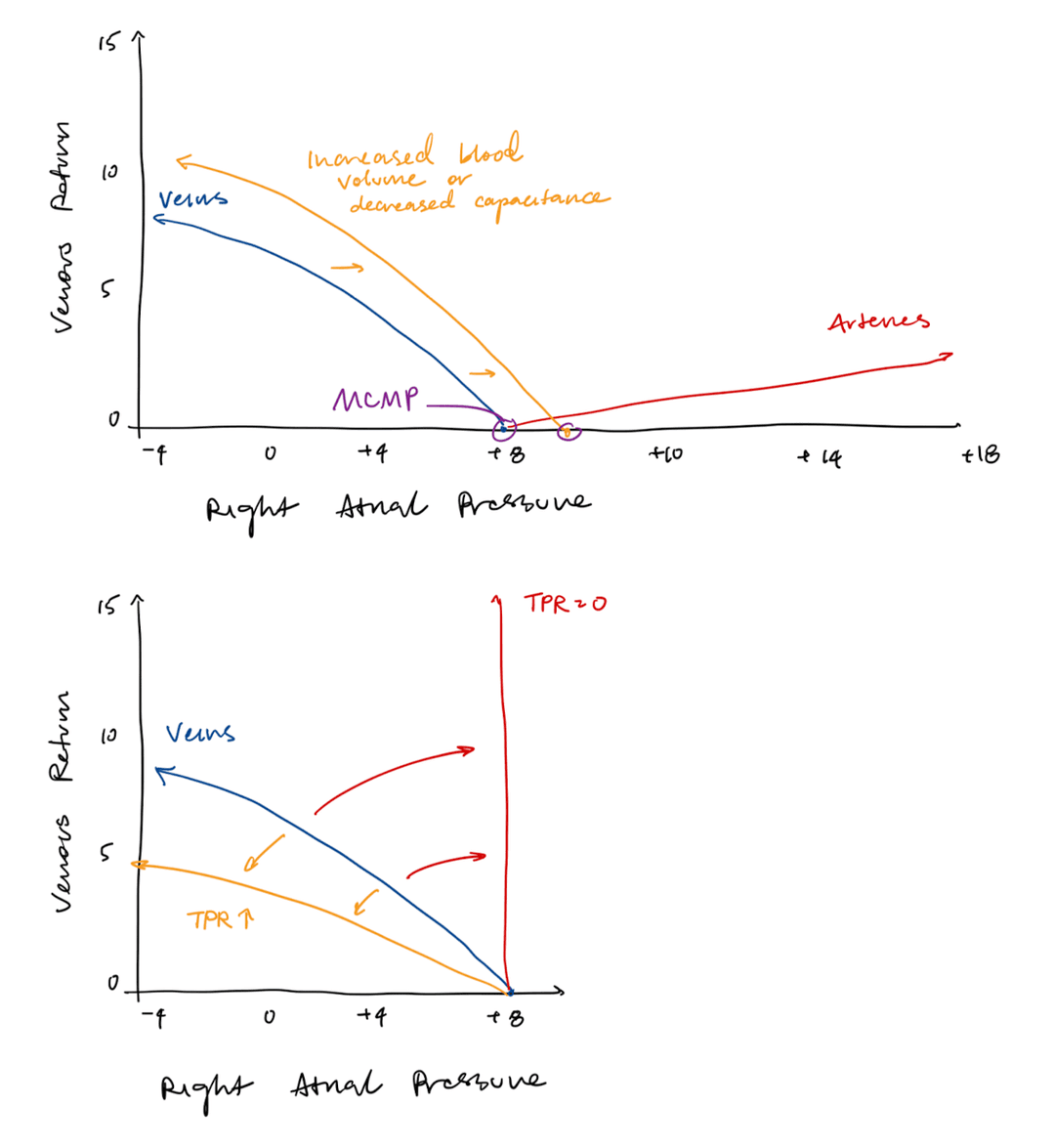
Cardiovascular Curve
At stead state, cardiac output is equal to venous return. Thus, the system should operate at the intersection of the cardio and vascular curves.
Exercise, heart failure, increased blood pressure, and hemorrhage (blood loss) can all affect the cardiac and vascular function curves. To determine how they affect the curves, consider:
- Direct effects
- Effect on blood pressure
- Sympathetic or parasympathetic response
Respiratory System
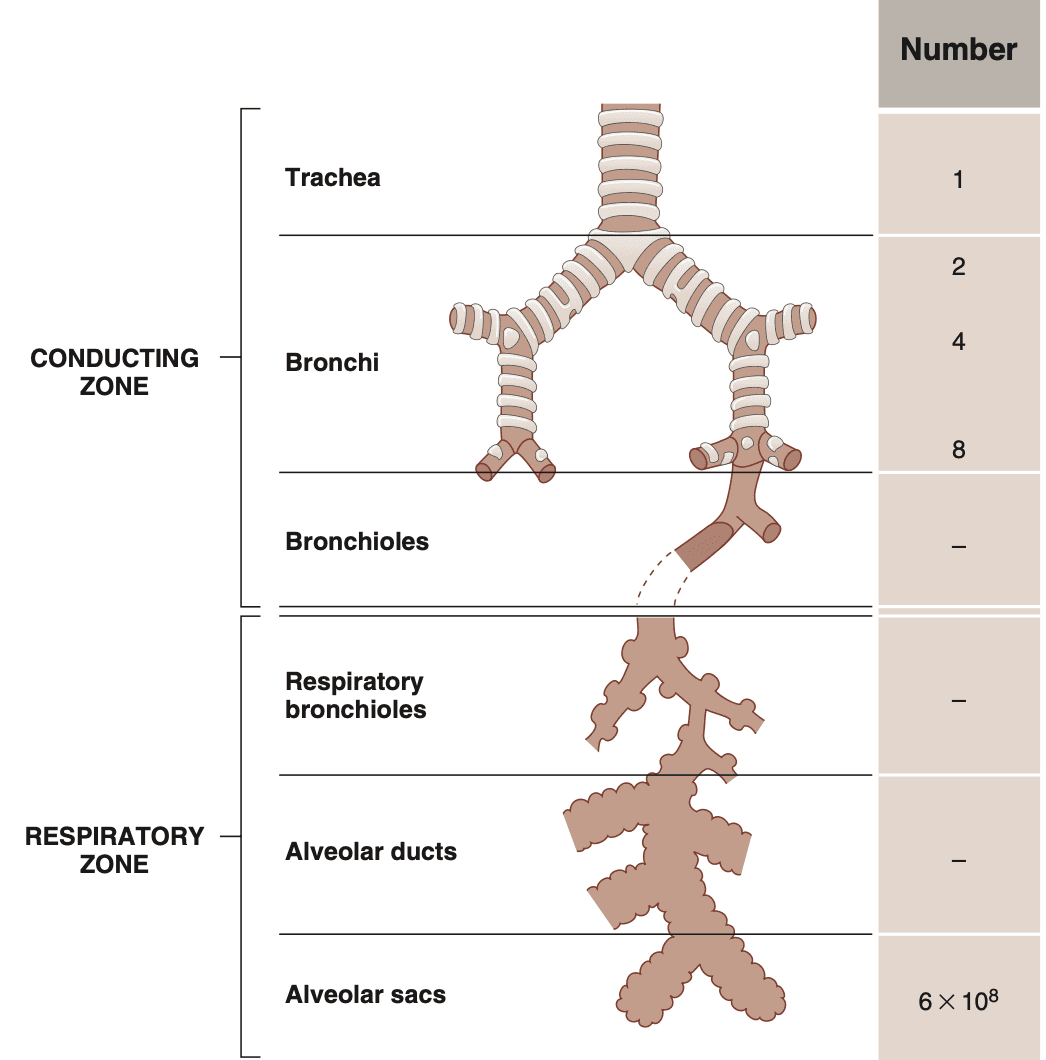
Structure of the Respiratory System
The respiratory system can be divided into two zones:
- The conducting zone brings in air
- The respiratory zone facilitates gas exchange
The conducting zone walls are lined with smooth muscles, which react to sympathetic and parasympathetic stimulation to eitehr dilate or constrict the airways.
The respiratory zone walls are lined with alveoli, where gas exchange occurs: O2 enters the pulmonary blood and CO2 leaves it.
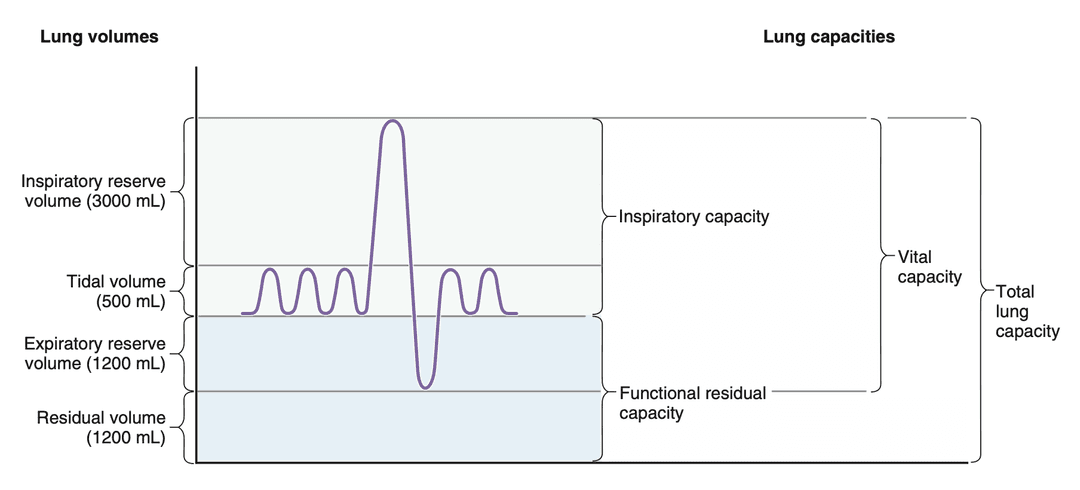
Lung Volume and Capacity
Lung volumes are measured with a spirometer.
First, the subject is asked to breathe normally, in order to measure the tidal volume (TV), which should be around 500 mL. Next, the subject is asked to take a maximal inspiration followed by a maximal expiration. The additional volume that can be inspired above the TV is called the inspiratory reserve volume (IRV), which should be around 3000 mL. The additional volume that can be expired below the TV is called the expiratory reserve volume (ERV), which should be around 1200 mL.
The volume of gas remaining in the lungs after a maximal forced expiration is the residual volume (RV), which is approximately 1200 mL and cannot be measured by the spirometer.
| Term | Definition |
|---|---|
| Tidal volume (TV) | Volume of air inhaled or exhaled during normal breathing |
| Inspiratory reserve volume (IRV) | Additional volume of air that can be inhaled |
| Expiratory reserve volume (ERV) | Additional volume of air that can be exhaled |
| Residual volume (RV) | Volume of air remaining in the lungs after maximum exhalation |
Lung capacities are combinations of lung volumes. For example, the inspiratory capacity (IC) is the maximum volume of air that can be inhaled after a normal exhalation, which is the sum of TV and IRV. The functional residual capacity (FRC) is the volume of air remaining in the lungs after a normal exhalation, which is the sum of ERV and RV. The vital capacity (VC) is the maximum amount of air that can be exhaled, and is composed of the IC plus the ERV. Finally, the total lung capacity (TLC) includes all of the lung volumes.
Because RV cannot be measured by spirometry, FRC and TLC also cannot be measured by spirometry.
| Term | Definition |
|---|---|
| Inspiratory capacity (IC) | Maximum volume of air that can be inhaled |
| Vital capacity (VC) | Maximum amount of air that can be exhaled |
| Functional residual capacity (FRC) | Volume of air remaining in the lung normal exhalation |
| Total lung capacity (TLC) | Maximum amount of air the lungs can hold |
Dead Space
Dead space is the volume of air that does not participate in gas exchange. For example, with a TV of 500 mL, around 150 mL fills the anatomic dead space, and 350 mL fills the alveoli. Dead space air is also the first air expired.
Physiologic dead space can differ from anatomic dead space when there is a defect in the ventilation-perfusion ratio. In that case, it can be calculated this equation:
Where:
- is dead space (mL)
- is tidal volume (mL)
- is arterial partial pressure of CO2
- is expired partial pressure of CO2
In other words, dead space is tidal volume multiplied by some fraction.
Ventilation Rates
Ventilation rate is the rate of air movement in the respiratory system. It can be expressed either as the minute ventilation, which is the rate of air movement into and out of the lungs.
It can also be expressed as alveolar ventilation, which is the rate of air movement into and out of the alveoli. Alveolar ventilation corrects for physiologic dead space.
Where;
- is alveolar ventilation (mL/min)
- is tidal volume (mL)
- is dead space volume (mL)
- is breathing frequency (breaths/min)
Mechanics of Breathing
The lungs tend to collapse inward due to their elasticity. The chest tends to expand outward. The pleural cavity between them maintains a negative pressure, which keeps the lungs inflated and allows these opposing forces to stay in balance.
Surface tension in the alveoli further promotes collapse, but the lungs produce pulmonary surfactant to reduce this tension. By lowering surface tension, surfactant increases lung compliance (capacitance), making it easier for the lungs to expand during breathing.
During inhalation, the diaphragm and intercostal muscles contract, expanding the thoracic cavity and making pleural pressure more negative. This decrease in pressure draws air into the lungs. During exhalation, these muscles relax, the thoracic cavity decreases in size, pleural pressure becomes less negative, and air flows out of the lungs.
In a pneumothorax, air enters the pleural space and disrupts the negative pressure. As a result, the lung collapses inward and can no longer expand properly.
Question: The cellular respiration equation has a 1:1 ratio of oxygen to carbon dioxide. Why do we breathe in more oxygen but breathe out less carbon dioxide?
The 1:1 ratio of O2 to CO2 applies only to glucose metabolism. In reality, the body also metabolizes fats and proteins, which contain less oxygen than carbohydrates. As a result, their oxidation requires more O2 per CO2 produced, so we consume more oxygen than the carbon dioxide we exhale.
This is captured by the respiratory exchange ratio, or the ratio of carbon dioxide produced to oxygen consumed. The average respiratory exchange ratio for a mixed diet is around 0.8.
Lung Conditions
Normal lung. PO2 in the alveoli and blood are around 100 mmHg.
Emphysema. Elastic fibers holding the small airways open are damaged, leading to difficulty breathing out. Additionally, the alveoli are damaged, so there is less surface area for gas exchange. This causes a decrease in PO2 in the blood.
Pulmonary edema. There is fluid in the alveoli, which increases the diffusion distance for gas exchange. This causes a decrease in PO2 in the blood.
Fibrotic lung disease. The alveoli are thickened, so there is a longer diffusion distance for gas exchange. This causes a decrease in PO2 in the blood.
Asthma. Increased resistance in the airways causes a decrease in airflow, which can lead to a decrease in PO2 in the alveoli and blood.
Ventilation-Perfusion Ratio
Ventilation (V) is the air reaching the alveoli per minute, and perfusion (Q) is the blood reaching the alveoli per minute. The V/Q ratio is around 0.8 in healthy individuals.
The body works to match ventilation and perfusion. When ventilation increases, perfusion also increases to maintain V/Q.
It also directs airflow and blood flow to optimize gas exchange. For example, the bottom of the lungs receives more blood due to gravity, so airways there dilate to increase ventilation.
Alveolar Gas Equations
The arterial partial pressure of oxygen and carbon dioxide can be calculated using the alveolar gas equations:
Where:
- is arterial partial pressure of CO2
- is alveolar partial pressure of CO2
- is barometric pressure
- is water vapor pressure
- is rate of CO2 production (mL/min)
- is alveolar ventilation (mL/min)
Note that we assume . This usually holds up unless diffusion between alveolar air and capillaries is limited.
Where:
- is arterial partial pressure of O2
- is alveolar partial pressure of O2
- is fraction of O2 in the alveoli
- is barometric pressure
- is water vapor pressure
- is rate of O2 production (mL/min)
- is alveolar ventilation (mL/min)
We expect to be around 40 mm Hg and to be around 100 mm Hg.
Oxygen Transport in Blood
In room air, oxygen and carbon dioxide act exactly the same. However, oxygen dissolves very poorly—it exerts 20x more partial pressure dissolved in water than CO2. This makes it difficult for blood to dissolve enough oxygen. Thus, most oxygen is actually bounded to hemoglobin.
The breakdown is:
- 1.5% dissolved in plasma
- 98.5% bound to hemoglobin
The oxygen-hemoglobin dissociation curve describes how the partial pressure of oxygen is related to the percent O2 saturation of hemoglobin. The curve is sigmoidal due to change in binding affinity and saturation. When one oxygen binds to hemoglobin, it increases the affinity of the remaining sites for oxygen.
When the blood leaves the lungs, the PO2 is around 100 mmHg and the hemoglobin is almost completely saturated. When the blood reaches the tissues, the PO2 level drops to around 40 mmHg, and the hemoglobin is around 75% saturated. This means that around 25% of the oxygen bound to hemoglobin is released to the tissues.
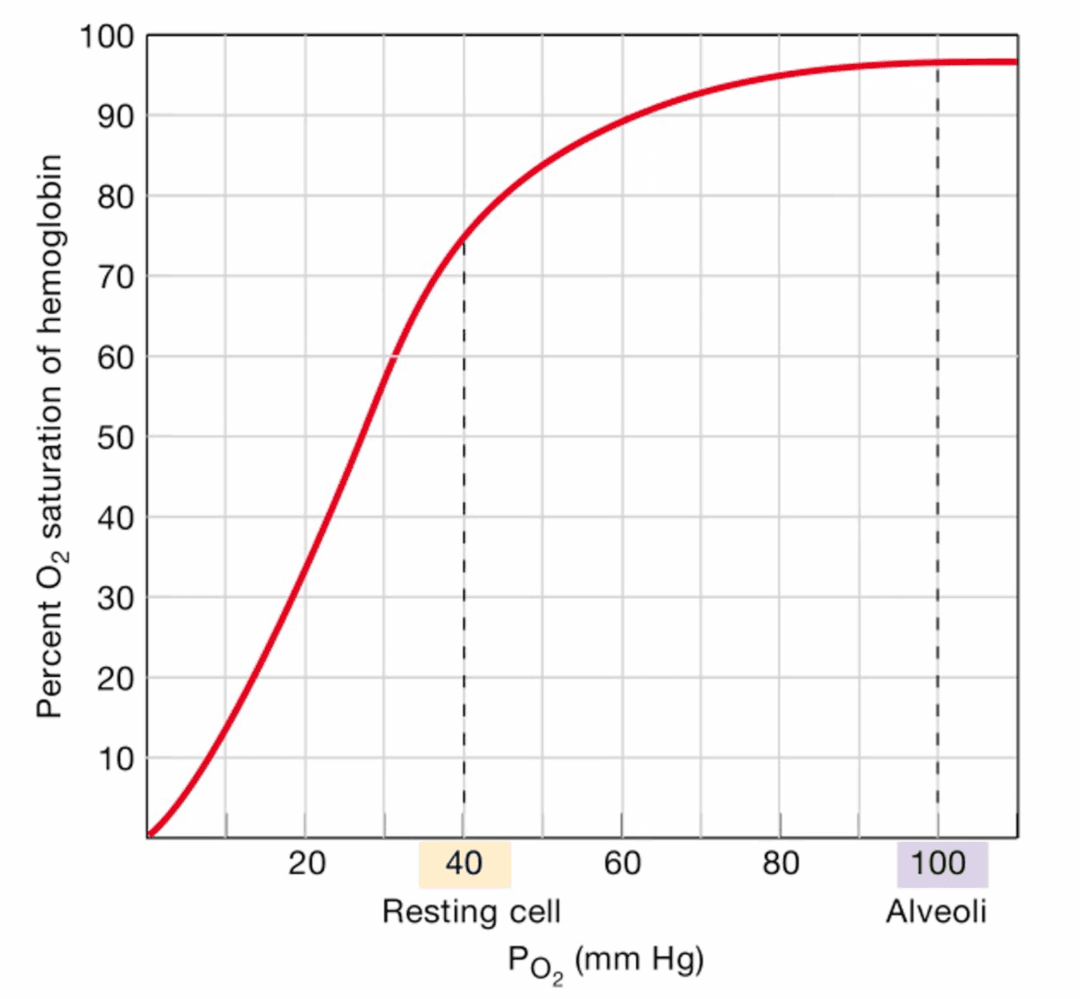
The Bohr effect states that the affinity of hemoglobin for oxygen decreases with
- Increased temperature
- Increased
- Increased [H+] (or decreased pH)
The Fick principle states that the rate of oxygen consumption by the tissues is equal to the product of cardiac output and the difference in oxygen content between arterial and venous blood:
Where:
- is the rate of oxygen consumption by the tissues (mL/min)
- is the cardiac output (mL blood/min)
- is the concentration of oxygen in arterial blood (mL O2/mL blood)
- is the concentration of oxygen in venous blood (mL O2/mL blood)
If the tissue is very metabolically active, then it will consume more PO2, leading to a lower point on the oxygen-hemoglobin dissociation curve. In addition, it will have a higher temperature and higher , leading to a Bohr shift to the right. Both of these effects will cause more oxygen to be released from hemoglobin, resulting in a lower concentration of . Thus, more oxygen will be delivered to the tissue. Note that changing is very difficult, but changing is much easier.
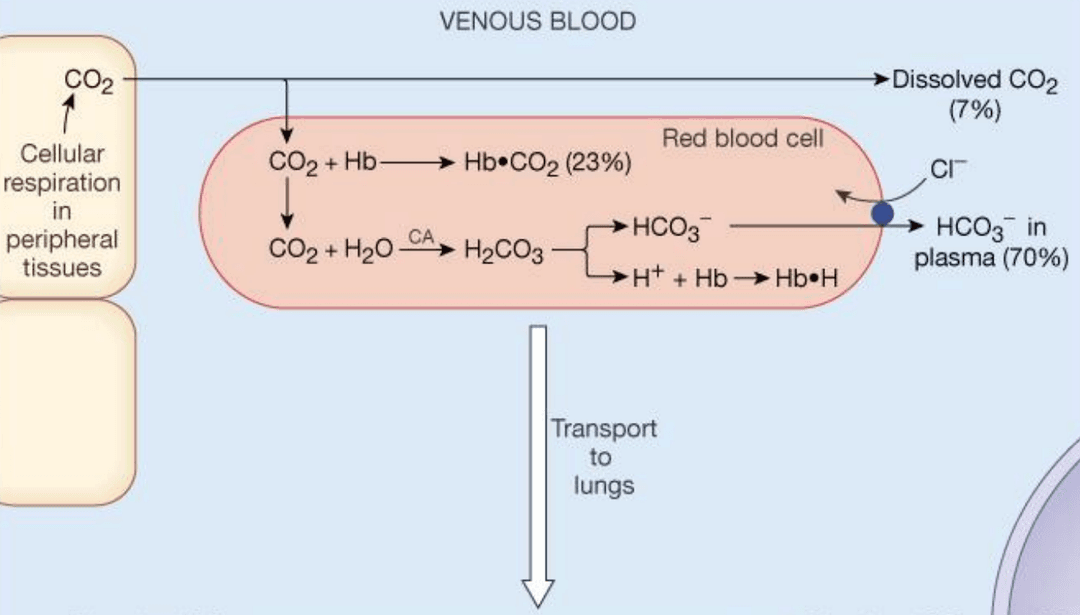
Carbon Dioxide Transport in Blood
Carbon dioxide is transported in the blood in three forms:
- 5% dissolved in plasma
- 10% bound to hemoglobin
- 85% as bicarbonate (HCO3-)
CO2 is converted to bicarbonate by the enzyme carbonic anhydrase in red blood cells. The reaction is:
By Le Chatelier's principle, when more CO2 is produced by the tissues, the reaction shifts to the right, producing more H+ and HCO3-. The H+ binds to hemoglobin, buffering the acidity. This also decreases hemoglobin's affinity for oxygen, causing more oxygen to be released to the tissues.
Control of Breathing
The brain monitors PO2, PCO2, and pH through chemoreceptors in the carotid bodies and aortic bodies. When PO2 decreases, PCO2 increases, or pH decreases, the brain increases ventilation to restore normal levels.
Rising CO2/falling pH is the first to regulate the 'urge to breathe'.
Renal System
The kidneys have two primary tasks:
- They regulate fluid volume and composition by varying the excretion of solutions and water
- They secrete three hormones: renin, erythropoietin, and 1,25-dihydroxycholecalciferol
Renal Anatomy
The kidneys have three main regions:
- The cortex is the outer region
- The medulla is the central region
- The papilla is the innermost and empties urine into the ureter, which is eventually transported to the bladder
The functional units of the kidneys are nephrons. Each kidney contains around 1 million nephrons. A nephron consists of a glomerular capillary network (glomerulus) surrounded by Bowman's space, as well as a renal tubule adjacent to peritubular capillaries.
Blood enters the kidney through the renal artery, which branches down to afferent arterioles that deliver blood to the glomerular capillaries. In the glomerulus, part of the plasma is filtered across the filtration barrier into Bowman’s space; this filtrate then flows into the proximal tubule. The blood that is not filtered leaves through the efferent arteriole, enters the peritubular capillaries, and eventually returns to the renal vein.
The renal tubule, which is lined with epithelial cells and serve the functions of reabsorption and secretion. The segments are called:
- Proximal tubule
- Loop of Henle
- Thick ascending limb
- Distal tubule
- Collecting ducts
Renal Blood Flow
The kidneys receive about 25% of the cardiac output, which is among the highest of all the organ systems.
The renal plasma flow (RPF) represents the amount of plasma flows through the renal system per minute and is typically around 500 mL. However, not all plasma is actually filtered across the glomerulus. The glomerular filtration rate (GFR) represents the amount of plasma is filtered across the glomerulus into Bowman's space per minute, and is only around 150 mL. The rest of the blood directly flows through the peritubular capillaries. Of the 150 mL, around 124 mL is reabsorbed; only 1 mL is actually excreted as urine.
The filtration fraction (FF) is equal to GFR divided by RPF, and it tends to be around 0.3.
Renal blood flow is the total volume of blood flowing to the kidneys per minute.
RBF is directly proportional to the pressure gradient between the renal artery and the renal vein, and it is inversely proportional to the resistance. The main mechanism for changing blood flow is by changing resistance through:
- Sympathetic system activates alpha-1 receptors, causing vasoconstriction mainly on the afferent arterioles
- Angiotensin-converting enzyme (ACE) activates angiotensin II, causing vasoconstriction mainly in the efferent arterioles (at low levels) or both arterioles (at high levels)
- Atrial natriuretic peptide (ANP) dilate afferent arterioles and constrict efferent arterioles, increasing GFR.
One tricky part is the difference between RBF and GFR. When the afferent arteriole is constricted, less fluid enters the renal system, decreasing both RBF and GFR, as expected. When the efferent arteriole is constricted, less fluid can move through the system, again decreasing RBF. However, the constriction also increases hydrostatic pressure in the glomerulus, leading to more fluid being pushed into Bowman's space, increasing GFR.
| Change | RBF | GFR | FF |
|---|---|---|---|
| Afferent dilation | Increase | Increase | No change |
| Afferent constriction | Decrease | Decrease | No change |
| Efferent dilation | Increase | Decrease | Decrease |
| Efferent constriction | Decrease | Increase | Increase |
There are also autoregulation mechanisms, including:
- Renal baroreceptors detect stretch in blood vesels, producing a myogenic responses which activate stretch-activated calcium channels and contract the muscle, causing an increased resistance to blood flow
- The macula densa in the juxtaglomerular apparatus in the distal tubule senses increased load and responds by secreting a substance that constricts afferent arterioles (via paracrines)
Renal Clearance
The clearance of a substance is the volume of blood that is completely cleared of a specific substance by the kidneys per unit of time. The higher the clearance, the more plasma that is cleared of the substance.
Clearance multiplied by the plasma concentration of the substance is equal to the amount of urinary excretion.
Where:
- is clearance (mL/min)
- is plasma concentration of the substance (mg/mL)
- is urine concentration of the substance (mg/mL)
- is urine flow rate (mL/min)
If the clearance of a substance is less than GFR, there is net reabsorption of that substance along the nephron. If the clearance of a substance is greater than the GFR, then there is net secretion along the nephron.
Inulin is a substance whose clearance is exactly equal to GFR. It is freely filtered across the glomerular capillaries, and is neither reabsorbed nor secreted. In other words, the amount of inulin filtered will be exactly equal to the amount of inulin excreted. Because of these properties, inulin is used as a glomerular marker.
The clearance of any substance can be compared with the clearance of inulin and is expressed as the clearance ratio. If , then the clearance is x is lower than the clearance of inulin. Either the substance is not filtered from the plasma, or it is filtered but subsequently reabsorbed. This includes albumin, Na+, Cl-, HCO3-, phosphate, urea, glucose, and amino acids.
If , then the clearance of x is higher than the clearance of inulin. The substance is filtered and secreted. This includes organic acids and bases, and sometimes K+.
Measurement of GFR, RPF and RBF
As noted above, GFR can be measured using the clearance of inulin:
RPF can be estimated from the clearance of para-aminohippuric acid (PAH). The main idea is that the amount of PAH entering the kidney () should be equal to the amount of PAH leaving the kidney (). This gives us the equation:
Where:
- RPF is renal plasma flow (mL/min)
- is the concentration of PAH in urine (mg/mL)
- is the urine flow rate (mL/min)
- is the concentration of PAH in the renal artery (mg/mL)
- is the concentration of PAH in the renal vein (mg/mL)
This is can be hard to sample, so some simplifying assumptions we can make is that is equal to zero, and equals the PAH concentration in any peripheral vein.
RBF can be calculated from RPF and hematocrit with the equation . Remember that hematocrit is the fraction of blood volume that is occupied by red blood cells, so 1 − hematocrit is the fraction of blood volume that is occupied by plasma.
Regulation of Water and Solutes
Antidiuretic hormone (ADH), or vassopressin, has three important effects:
- Increase water permeability in distal tubule and collecting ducts
- Increase activity of Na+-K+-2Cl- cotransporter in thick ascending limb
- Increases urea permability in collecting ducts They effect of water permeability is the most important; in essence, ADH tells the kidneys save more water.
Atriopeptin (ANP) is triggered by a low pressure sensing system in the atria. As blood volume increases, the atria stretches, causing more ANP to be produced. In the kidneys, it causes vasodilation of afferent arterioles, vasoconstriction of efferent arterioles, increased GFR, and decreased Na+ reabsorption in the late distal tubule and collecting ducts.
The job of the renin-angiotensin-aldosterone system (RAAS) is to raise blood pressure and increase blood volume. It is activated by low blood pressure and low blood volume.
- First, renin is released by the juxtaglomerular cells in the afferent arteriole. Renin cuts angiotensinogen (from the liver) into angiotensin I, which is then converted by ACE into angiotensis II.
- As discussed before, angiotensin II causes vasoconstriction, increasing resistance and blood pressure. It also stimulates the release of vasopressin and aldosterone, and increases thirst levels.
- Aldosterone increases Na⁺ reabsorption in the distal tubule and collecting duct, causing water to follow the Na+. It also stimulates A-type cells, causing more HCO3- and K+ to be reabsorbed. It also causes more H+ to be excreted, preventing acid from accumulating in the body.
Note that carbonic anhydrase is necessary for:
- Secretion of acid in the form of NH4+ in the proximal tubule
- Secretion of acid in the distal tubule
- Secretion of bicarbonate in the distal tubule
Free-Water Clearance
Free water is water that is not accompanied by solute. Positive free water clearance means that the kidney is excreting solute-free water, usually with low ADH and hyposmotic urine. Negative free water clearance means that the kidney is reabsorbing free water, usually with high ADH and hyperosmotic urine. All of the free water generated in the thick ascending limb and the early distal tubule is reabsorbed by the late distal tubule and collecting duct.
Acid-Base Physiology
The body human body maintains an arterial pH of 7.35.
Bicarbonate Buffer System
The bicarbonate buffer system is the main pH buffer system. First, CO2 can react with H2O to convert to H2CO3 (carbonic acid) when catalyzed by carbonic anhydrase. H2CO3 itself dissociates into H+ and HCO3 (bicarbonate). The H+ generated by this reaction must be buffered.
Here is how they buffer the pH:
- CO2 levels are controlled by the lungs
- Bicarbonate (HCO3) excretion is controlled by the kidneys
- To buffer acids, HCO3 reacts with H to form H2CO3, which then dissociates into CO2 to be exhaled by the lungs
- To buffer bases, H2CO3 dissociates into H and HCO3, which is put in the bicarbonate reserve in the kidneys
Henderson-Hasselbalch Equation
Let be the chemical equilibrium constant for a buffer base or acid:
By the Henderson-Hasselbalch equation:
Where:
- is the concentration of the base form
- is the concentration of the acid form
Therefore the pH of a buffered solution can be calculated with the following information: the pK of the buffer, the concentration of the base form of the buffer, and the concentration of the acid form of the buffer.
Strong acids will have a high equilibrium constant but low pKs, while weak acids will have low equilibrium constants but high pKs.
The bicarbonate system has an effective pKa of 6.1. For the blood to be at a pH of 7.35, physiologically there is a lot more bicarbonate than carbonic acid present in the blood.
Compensatory Responses
Compensation for acid-base disorders is either respiratory or renal. When the primary disorder is metabolic, compensation is respiratory. When the primary disorder is respiratory, compensation is renal (metabolic).
Alkalosis is a condition where arterial pH is too basic.
- Metabolic alkalosis is caused by increased HCO3- in the blood. This often happens in vomiting, when the acidic stomach contents are lost. It can be compensated by hypoventilation in the lungs, which increases CO2, which can be converted to carbonic acid.
- Respiratory alkalosis is caused by hyperventilation, which results in the loss of CO2. It can be compensated by decreased activity of acid secreting cells and decreased HCO3- reabsorption in the kidneys.
Acidosis is a condition where arterial pH is too acidic.
- Metabolic acidosis is caused by decreased HCO3- in the blood. It can be compensated by hyperventilation in the lungs, which will decrease CO2 and raise pH.
- Respiratory acidosis is caused by hypoventilation, which results in the retention of CO2. It can be compensated by increased H+ secretion and excretion of H+,and increased sythesis and reabsorption of new HCO3- in the kidneys.
Typical arterial blood gas values are 26 mM/L HCO3- and 40 mmHg PCO2.
Exercise Physiology
Physical units
Performance can be quantified in terms of physical units
- Force is in newtons, or kg-m/s^2
- Work is in joules, or N-m
- Power is in watts, or N-m/s
1 newton is the amount of force required to hold 102g up against Earth's gravity. 1 joule is the amount of work needed to lift 102g up one meter. 1 watt is the amount of power need to lift 102g up one meter in one second.
Calories are also a measurement of work. 1 KCal/min is around 70W.
Muscle Fibers
There are three types of muscle fibers:
- Slow-twitch is the least powerful
- Fast-twitch a is intermediate and fatigue-resistant
- Fast-twitch b is the most powerful but fatigable
There is great genetic variability in the distribution of muscle fibers between individuals. e.g. In the gastrocnemius (calf) muscle, elite distance runners have 90% ST and elite sprinters have 20% ST.
Energy Systems
- The phosphagen system (10s) uses stored ATP and phosphocreatine and produces energy extremely quickly
- The glycolytic system (1-2 min) breaks down carbohydrates through glycolysis, resulting in buildup of lactic acid
- The oxydative system (aerobic) (indefinite) metabolizes both carbs and fatty acids. It is very efficient, and fatigue-resistant, but also very slow
As the requirement for more power from muscle contraction increases, the recruitment order is as follows:
- Slow-twitch (aerobic)
- Fast-twitch a (aerobic/anaerobic)
- Fast-twitch b (anaerobic)
The old story about the anaerobic threshold is as follows: as exercise intensity increases, oxygen levels are insufficient to support aerobic pathways. This causes the body to fall to the phosphagen and glycolytic system. Pyruvate, the product of glycolysis, accumulate. Pyruvate gets converted to lactate, which accumulates as a "waste product" and enters the bloody. The buffering of lactate by HCO3- results in increased VCO2.
However, many subsequent studies do not support this hypothesis. Lactate is a resource that can also be used as fuel, be converted to glucose (in liver), and serve as a signalling molecule.
Instead, we see a switch from fat to carbohydrates:
- Slow twitch muscles switch from fat to carbs to generate more power.
- More fast twitch muscles are recruited, which exclusively use glycolysis
As glycogen stores are exhausted, one may enter excess post-exercise oxygen consumption (EPOC). There are two components to it.
- The fast component, or the alactacid oxygen debt, is the immediate oxygen used right after exercise to rapidly restore oxygen stores. It mainly includes reoxygenating myoglobin and restoring ATP and creatine phosphate stores.
- The slow component, or the lactaid oxygen debt, involves dealing with metabolizing lactic acid and returning metabolism to baseline.
Power Output
Minute ventilation is proportional to power output until after 400 Watts. This is because CO2 levels rise proportionally until a certain point, at which CO2 increases disproportionately. This is the point when muscles go anaerobic, and the buffering of lactic acid with HCO3- generates additional CO2.
Digestive System
The digestive system is composed of the gastrointestinal (GI) tract and salivary glands, liver, pancreas, and gallbladder.
Most organs in the digestive system are primarily composed of smooth muscles. The GI tract also has a mucous barrier for physical and chemical protection.
Gastrointestinal Anatomy
The GI tract includes:
- Mouth
- Pharynx
- Esophagus
- Stomach (pH 1-2)
- Small intestine
- Large intestine (including the colon)
- Rectum
- Anus
Note that the trachea (windpipe) and esophagus (food pipe) are adjacent to each other.
The appendix is attached to the first portion of the large intestine. The appendix in humans is useless, but koalas use them to ferment eucalyptus leaves and digest bacteria. Diarrhea occurs when sodium and water is pumped into the digestive system in order to rinse it out. This gets rid of both good and bad bacteria. People with healthy appendices can more quickly populate with good bacteria.
The GI tract is divided via sphincter muscles:
- Upper esophageal sphincter (between mouth and pharynx)
- Lower esophageal sphincter (between pharynx and stomach)
- Pyloric valve (between stomach and small intestine)
- Illeocecal valve (between small intestine and large intestine)
- Internal anal sphincter (smooth muscle)
- External anal sphincter (striated muscle)
The upper esophageal sphincter prevents air from entering the esophagus. It only relaxes during the swallowing reflex. The lower esophageal sphincter prevents the acidic stomach contents from coming up the esophagus (heartburn). It is relaxed during the vomiting reflex
The pyloric valve regulates the flow of stomach into the small intestine. The role of the small intestine is to extract all nutrients from the food before it enters the large intestine.
Stomach
The stomach and intestine consists of
- Endothelial cells
- Smooth muscle
- Neurons
- Glandular (duct) cells with exocrine, endocrine, or paracrine secretions function
The stomach digests food by secreting mucus, acid, and digestive enzymes
- Parietal cells secrete acid (HCI)
- Chief cells secrete digestive enzymes like pepsinogen and gastric lipase
- D cells release somatostatin in response to HCl, which basically inhibit gastic acid, gastrin, and pepsinogen
- ECL cells release histamine, which stimulates parietal cells to secrete more acid
The phases of digestion include the cephalic phase, the gastric phase, and the intestinal phase. During the intestinal phase, the enteric nervous system receives feedback. Then enterogastrones are released from enteroendocrine glands in the small intestine.
Examples include:
- Secretin is triggered by acid. It inhibits acid secretion and stimulates bicarbonate secretion from the pancreas
- Cholecystokinin (CCK) is triggered by fats. It inhibits acid secretion and stimulates the release of bile from the gallbladder. Bile emulsifies large bits of fat into smaller bits of fat. CCK also decreases feeding behavior.
- Gastric inhibitory peptide (GIP) is triggered by carbs. It inhibits acid secretion and stimulates pancreatic beta cells (for insulin release). Insulin is necessary for transporting sugar into muscle tissue and fat molecules. Excess carbs are stored as fat (glycogen) for long-term storage.
- Glucagon-like peptide (GLP-1 ) is triggered by carbs. It stimulates pancreatic beta cells (which secrete insulin) and alpha cells (which decrease glucagon).
CCK, GIP, and GLP-1 result in decreased feeding behavior.
Liver
The liver eliminates bilirubin and produces bile salts to emulsify fat. Bilirubin is a byproduct of the breakdown of hemoglobin. Bile salts emulsifies lipids, breaking large droplets into smaller droplets to increase surface area for metabolic activity.
Leptin, a hormone released from fat cells, can also activate the autonomic nervous system (sympathetic and parasympathetic tones) and decrease the release of glucose from the liver.
Pancreas
The pancreas is responsible for releasing insulin (alpha cells) and glucagon (beta cells). When insulin binds to receptors, it allows GLUT-4 transporters to be inserted into the membrane, allowing glucose to diffuse. Insulin also causes glucose levels to drop inside the cell, causing glucose to be transported into the cell. Conversely, glucagon raises blood glucose levels.
When sugar levels rise, insulin levels rise and glucagon levels drop.
Insulin:
- Increases glucose uptake in muscle cells, fat cells, and in the liver via GLUT-4 transporters
- Increases glycogen and protein synthesis in muscle cells
- Increases triglyceride synthesis in fat cells
- Increases glycogen and triglyceride synthesis in the liver
- Turns off ketone synthesis in the liver
Enterokinases in brush border of small intestine
- The pancreas secretes inactivate proenzymes into the lumen of the small intestine
- Among them, trypsinogen is cut into the shorter protein trypsin by brush border enzyme enteropeptidase
- Trypsin is activated by enteropeptidase and activates remaining proenzymes by cutting peptide bonds
- Proteins, carbs, and fats are now digested in small intestine by activated enzymes
Large Intestine
- Stories and dries feces
- Absorbs some nutrients (fiber)
- Host for "good" bacteria (microbiome)
- Gut-brain communication
Probiotics are live microorganisms that, when administered to a host, have health benefits. They can:
- Compete out harmful microorganisms
- Strengthen tight junctions
- Activate hormone signalling
Metabolism
Nutrients
Carbohydrates like polysaccharides, sucrose, lactose, and fiber are broken down into monosaccharides (glucose, galactose, fructose). Proteins, or polypeptides, are broken down into amino acids. Fats like triglycerides are broken down into monoglyceride and free fatty acids
Humans do not have cells that can break down cellulose (nutritionally called fiber), but there is bacteria in the large intestine that can break it down.
Absorptive State
Neurons metabolize glucose from blood. When glucose is not available, they can also metabolize ketones. All other cells metabolize fatty acids. When present, they can also metabolize ketones. Muscles can metabolize stored glycogen, which produces lactate and pyruvate.
When glucose levels are low, the liver releases glucose into the blood from stored glycogen and from glycerol, lactate, and pyruvate (through gluconeogenesis). They can also convert fatty acids to ketones.
The absorptive state is the period after a meal and has the following properties:
- Insulin levels are high
- Glucagon levels are low
- Many tissues use glucose as their main energy source
- The liver takes up glucose and stores energy as glycogen
- The liver stops producing new glucose via gluconeogenesis
- Ketone production is suppressed by insulin
Feeding behavior is controlled by:
- Increased levels of cholecytokinin (CCK)
- Stretch of stomach
- Rise in blood levels of glucose and insulin
Diabetes
Type 1 Diabetes
- Autoimmunity against beta cells from the pancreas
- Can be controlled with insulin therapy
- Symptoms include elevated sugar levels in blood and urine; increased eating and drinking; increased potassium levels
Type 2 Diabetes
- Receptors to insulin not as effective
- Can be controlled by diet, exercise, and insulin therapy
Pathophysiology
Hyperkalaemia
Hyperkalaemia is defined as high extracellular [K+] (> 5.5 mEq/L).
Its symptoms are:
- Fatigue and muscle weakness
- Arrhythmia (QRS widening, or longer ventricle depolarization)
- Bradycardia (slow resting heart rate)
- Paresthesia (abnormal sensory signals like pins or needles)
Hyperkalemia can be treated by injecting insulin or glucose. Insulin increases uptake of solute that bring Na+ into cells. This necessitates the pumping of more Na+ out of cells in exchange with K+ going into cells, decreasing extracellular K+. Insulin also stimulates the Na+/K+ ATPase pump, which results in more K+ being pumped into cells and a lowering of extracellular K+.
Congestive Heart Failure
Congestive heart failure is defined as decreased contractility of the heart. Kidneys retain Na+, Cl-, and H2O to increase mean circulatory filling pressure to restore stroke volume and cardiac output. This leads to fluid accumulation in the feet, lungs, and arms.
Causes include:
- Coronary artery disease
- Heart attack
- Infections, drug abuse
- Overworking the heart (high blood pressure, insufficient valves)
Treatments include:
- Exercise
- ACE inhibitors or angiotensin II inhibitors (reduce vassopressin)
- Beta blockers (if high blood pressure)
- Diuretics, restrict salt and water
Types of diuretics:
- Carbonic anhydrase inhibitors: inhibit CA in proximal tubule
- Loop diuretics: inhibit Na+/K+/2Cl- transporters in thick ascending loop
- Thiazide diuretics: block Na+/Cl- cotransporter in distal tubule
- Potassium sparing diuretcs: act on collecting duct
IBD and IBS
Inflammatory bowel disease (IBD) is an autoimmune disease causing inflammation and damage to the GI tract. Treatment includes immunosuppressant drugs and pain medication.
Irritable bowel syndrome causes no permanent damage. Treatment includes diet and pain medication.